Lab closed in 2005.
Axon Guidance and Target Recognition
It is a fascinating but daunting problem: How do just a few thousand genes direct the assembly of neuronal circuits with such staggering complexity as those of the human brain? We chose to tackle the fly first. It’s not that the fly’s nervous system is any less challenging, but at least we have a powerful set of genetic tools to work with. And what we learn about the development of the fly’s nervous system may provide new insights into our own. After all, it seems that there’s a bit of a fly in all of us.
Introduction
Neuronal circuits are formed as each neuron sends out axons and dendrites to find, recognize and connect with the appropriate target cells. Finding the target is the task of the growth cone, a highly motile and exquisitely sensitive structure at the tip of each axon and dendrite. Growth cones detect various guidance cues in the extracellular environment, and somehow manage to extract from this information the correct route towards their target. By studying this process in Drosophila, we hope to find out what these cues are, how growth cones detect them, and how each growth cone knows which cues it should follow and which it should ignore. Genetics is a powerful tool in this endeavor. Mutations that disrupt neuronal connectivity can lead us to the genes that encode the guidance cues themselves, or to the molecules that growth cones use to detect and respond to these cues. And, by examining how these molecules work in vitro and in vivo, we can begin to explore the molecular, cellular and developmental mechanisms that ensure that each growth cone responds to the right cues at the right time, and ignores the rest. We are focussing on two different systems: the ventral nerve cord of the embryo, and the adult visual system.
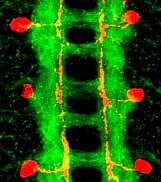
The ventral nerve cord of a Drosophila embryo. Axons are stained green. They form two main longitudinal tracts that run each side of the midline, and a series of commissures that cross the midline to connect the two symmetric halves of the nervous system. One neuron in each hemisegment is stained red. Its axon grows towards but not across the midline, before turning to run alongside it.
Axon guidance in the ventral nerve cord
n bilaterally symmetric nervous systems, such as our own and the fly’s, axons in the CNS must decide whether or not to grow across the midline. Our recent work has revealed how this decision is controlled in the Drosophila nerve cord (see figure below). Crossing and non-crossing axons differ in their sensitivity to the midline repellent Slit. Both commissural axons (which cross) and ipsilateral axons (which don’t) express the Slit receptor Roundabout (Robo for short). In ipsilateral axons, Robo is inserted into the growth cone, making them sensitive to the repulsive activity of Slit. But in commissural neurons, Robo is not delivered to the growth cone until after it has crossed the midline. So these axons are able to cross, but only once. Before crossing, an intracellular sorting receptor called Commissureless (Comm) collects the newly synthesized Robo protein at the Golgi and delivers it to lysosomes, where it is degraded. Comm is normally active only in commissural neurons as their axons first grow toward and across the midline. If it is made inactive (by a mutation), no axon can cross the midline, resulting in the commissureless phenotype from which the gene got its name. In ipsilateral neurons, and post-crossing commissural neurons, Comm is normally inactive. But by activating Comm in these neurons, we can force them to cross (or recross). This defines Comm as a simple genetic switch to control midline crossing (see figure below). We are currently trying to find out how this switch is turned ON and OFF, and how Comm selects Robo and sorts it to lysosomes.
Crossing the midline. In the Drosophila CNS, some axons cross the midline (like the red axons in A), but others do not (B). Those that do cross, called commissural axons, cross only once. Comm functions as a switch to control this decision (C). Comm is ON in commissural neurons as they cross, but OFF in ipsilateral neurons and post-crossing commissural neurons. Comm regulates the sensitivity of axons to the midline repellent Slit (yellow in C). It does this by controlling the intracellular trafficking of Robo (blue), the Slit receptor.
Axon guidance in the visual system
Flies have excellent vision. This rests in part on the extraordinarily precise connections established between photoreceptors in the eye and their targets in the brain. As a first step in determining how these connections are established, we screened though more than 32,000 mutant lines to find some 200 mutants with abnormal connectivity patterns. These mutations define about 50 different genes, 31 of which we have now identified. These genes encode cell surface receptors and signaling molecules, as well as factors involved in gene transcription, axonal transport, and membrane trafficking. One of them encodes a 7-transmembrane cadherin called Flamingo. Flamingo is expressed on many different photoreceptor axons and their target cells in the brain (see figure below). It is needed for one class of photoreceptors (R8s) to select their correct targets in a specific layer of the brain. We continue to look for the remaining genes, and are beginning to piece together the molecular pathways and processes that underlie this exceptional example of neuronal engineering.
Photoreceptor axon targeting. Photoreceptor axons (green) extend from the eye disc, through the optic stalk, and into the optic lobe. Some axons terminate in the lamina (arrow). Others, including R8 axons, terminate in the medulla (arrowhead), where they form synapses with medulla neurons (red). The Flamingo protein (blue) is required for correct targeting of R8 axons.
Selected Recent Publications
Demir, E., and Dickson, B. J. (2005). fruitless Splicing Specifies Male Courtship Behavior in Drosophila. Cell 121, 785-94.
Keleman, K., Ribeiro, C., and Dickson, B. J. (2005). Comm function in commissural axon guidance: cell-autonomous sorting of Robo in vivo. Nat Neurosci 8, 156-63. Epub 2005 Jan 16.
Stockinger, P., Kvitsiani, D., Rotkopf, S., Tirian, L., and Dickson, B. J. (2005). Neural Circuitry that Governs Drosophila Male Courtship Behavior. Cell 121, 795-807.
Keleman, K., Rajagopalan, S., Cleppien, D., Teis, D., Paiha, K., Huber, L. A., Technau, G. M., and Dickson, B. J. (2002). Comm sorts robo to control axon guidance at the Drosophila midline. Cell 110, 415-27.
Hakeda-Suzuki, S., Ng, J., Tzu, J., Dietzl, G., Sun, Y., Harms, M., Nardine, T., Luo, L., and Dickson, B. J. (2002). Rac function and regulation during Drosophila development. Nature 416, 438-42.